
Аннотация
Семьи диких медоносных пчел — как истинно дикие (на деревьях и в зданиях), так и имитирующие дикие (в небольших ульях) — изучались с целью определения их жизненных особенностей и выяснения, изменились ли они после заражения семей Varroa destructor. Большинство семей (97%) переживают лето, но только 23% семей-основательниц (первогодок) и 84% семей второго года переживают зиму. Средняя продолжительность жизни устоявшихся семей составляет 5-6 лет, и в большинстве из них (87%) каждое лето происходит смена маток (вероятно, роение). Популяционная модель показывает, что эти особенности жизненного цикла приводят к формированию стабильной популяции семей. Примечательно, что набор признаков жизненного цикла семей, обнаруженных в 2010-х годах (с V. destructor), совпадает с набором признаков, обнаруженным в 1970-х годах (без V. destructor). Представляется вероятным, что дикие семьи, обитающие в окрестностях Итаки (штат Нью-Йорк), обладают защитными свойствами против V. destructor, которые не требуют больших затрат.
Введение
Начиная с 1952 г. в Европе и с 1987 г. в Северной Америке семьи ульевых медоносных пчёл (Apis mellifera) стали заражаться эктопаразитическим клещом (Varroa destructor) (обзор Locke 2016). Исследователи обнаружили, что если семью медоносных пчел европейского происхождения не обработать от V. destructor, то через год-два популяция клещей в семье резко возрастет, титры вирусов в пчёлах подскочат, и семья погибнет (Korpela et al. 1992; Fries et al. 2006). Поэтому было принято считать, что дикие семьи медоносных пчёл европейского происхождения, обитающие в Европе и Северной Америке, погибли (Moritz et al. 2007; Potts et al. 2010).
Однако в середине 2000-х годов из Европы и Северной Америки стали поступать сообщения о популяциях диких семей европейских медоносных пчел, которые выживают, несмотря на заражение V. destructor: Швеции (Fries et al. 2006), Франции (Le Conte et al. 2007) и США (Seeley 2007). Сбалансированные отношения между медоносными пчелами и V. destructor ожидаются там, где большинство семей живет в дикой природе, поскольку в этих местах должен происходить сильный естественный отбор на устойчивых к клещу пчёл, а также, возможно, на низковирулентных патогенов и паразитов. Ожидание сильного естественного отбора на пчёлах было недавно подтверждено исследованием (Михеев и др., 2015) диких медоносных пчёл, обитающих в окрестностях Итаки (штат Нью-Йорк). В нём использовалось секвенирование всего генома пчёл, собранных из диких семей в 1977 г. (до появления V. destructor) и в 2011 г. (после появления V. destructor), и было обнаружено, что 232 ядерных гена, разбросанных по всему геному, подверглись сильному отбору между датами отбора. Исследование также показало, что в период с 1977 по 2011 год произошел обвал эффективной численности популяции этих пчёл, что, вероятно, явилось результатом массовой гибели семей после появления V. destructor в середине 1990-х годов. Однако численность переписной популяции восстановилась (Seeley 2007).
В данной работе от генетики переходят к экологии диких семей, обитающих в окрестностях Итаки (штат Нью-Йорк). В ней сообщается о 7-летнем исследовании (2010-2016 гг.) выживаемости этой популяции, несмотря на то, что все семьи заражены V. destructor. Исследование, проведенное 40 лет назад (1974-1977 гг.), показало, что дикие семьи, обитающие в окрестностях Итаки, обладают определенным набором особенностей жизненного цикла: медленное развитие, большая продолжительность жизни и малое, но дорогостоящее потомство (Seeley 1978). В настоящем исследовании выясняется, изменились ли особенности жизненного цикла этих семей с 1970-х годов, поскольку теперь им приходится вкладывать средства в иммунологическую, поведенческую и другие виды защиты от V. destructor и вирусов, переносчиками которых является этот клещ. Предположительно, затраты на эти дополнительные средства защиты компенсируются другими компонентами приспособленности, о чем говорится в работах Minchella (1985), Sheldon and Verhulst (1996) и Schmid-Hempel (2003), поэтому, возможно, у диких семей замедлилось развитие, сократилась продолжительность жизни или уменьшилось количество (или снизилась стоимость) потомства. Однако возможен и другой вариант: дикие семьи имеют малозатратные средства защиты от клещей и вирусов. Если это так, то параметры их жизненного цикла не изменятся. Чтобы выяснить, к какому сценарию относятся дикие семьи, обитающие в окрестностях Итаки (штат Нью-Йорк), я исследовал 33 гнездовых участка (на деревьях и в зданиях), занятых дикими семьями, и 22 улья, занятых имитацией диких семей.
2 Материалы и методы
2.1 Территория исследования и родословная пчёл
В данном исследовании использовались семьи, обитающие в сильно заросшей лесом местности к югу от Итаки (штат Нью-Йорк, США) (42° 26′ 36″ с.ш. 76° 30′ 0″ з.д.). Этот регион был заселен в начале 1800-х годов, и до конца 1800-х годов здесь велось преимущественно сельское хозяйство. В настоящее время сельское хозяйство прекращено, а на больших площадях, которые когда-то обрабатывались, выросли леса — от старопахотных сукцессионных до зрелых. Климат здесь умеренно-континентальный: лето короткое и теплое (температура редко превышает 32°С), зима длинная и холодная (температура часто достигает -18°С). Родословная диких семей проанализирована (Михеев и др., 2015). Как старые (1977 г.), так и современные (2011 г.) популяции являются в основном потомками центрально-европейских подвидов (Apis mellifera carnica и A. m. ligustica), но также имеют значительное число предков западноевропейских (A. m. mellifera) и среднеазиатских (A. m. caucasica) подвидов.
2.2 Поиск диких семей
Было прослежено 33 гнездовых участка диких семей, 14 из которых были найдены в результате охоты на пчел (Seeley 2016), а 19 — случайно. Как и ранее (Seeley 1978), я выделял две категории семей: семьи-основатели и сформировавшиеся семьи. Семьи-основатели — это те, которые еще не пережили первую зиму, а сформировавшиеся семьи — те, которые пережили. Семья считалась основанной или сформировавшейся только в том случае, если владелец дерева или здания, на котором находится семья, сообщал, что она поселилась на этом месте летом (следовательно, это семья-основатель) или жила там предыдущим летом и пережила предыдущую зиму (следовательно, это сформировавшаяся семья). Все остальные семьи были отнесены к категории «неоднозначных». Основательная или неоднозначная семья, пережившая зиму, следующей весной переводилась в категорию сформировавшихся семей.
2.3 Осмотр гнездовых участков и статистический анализ
Я осматривал каждое гнездо три раза в год: весной, летом и осенью. Весенние проверки проводились до 10 мая, т.е. до начала основного роевого сезона. Летние проверки проводились после 20 июля, т.е. после окончания основного роевого сезона. Осенние проверки проводились после 20 сентября, т.е. после окончания сезона вторичного роения. В районе Итаки 80% роев образуется в период с 15 мая по 15 июля и 20% — с 15 августа по 15 сентября (Fell et al. 1977). Летающие пчелы у входа в гнездо не являются доказательством наличия живой семьи; это могут быть воровки или разведчики, обследующие гнездо погибшей семьи. Критерием живой семьи для меня были пчёлы, прилетающие в гнездо с пыльцой. Я продолжал осматривать гнезда, в которых погибли семьи, для измерения частоты повторного заселения, за исключением пяти мест, которые были повреждены (например, два дупла в стенах домика были вскрыты черным медведем Ursus americanus).
Для проверки различий между летом и зимой в вероятности выживания семей были объединены данные о количестве семей, выживших или погибших в каждом сезоне (летом или зимой) за все 7 лет, в таблице с осями лето/зима, основатель/учреждение, выжил/погиб (см. Приложение 1, табл. IV). Эти данные были проанализированы отдельно для летних и зимних семей с проверкой однородности доли выживших семей между основателями и укоренившимися семьями с помощью точного теста Фишера. Этот тест предполагает независимость выживаемости семей по годам. Это предположение подтверждается значительными различиями между годами в погодных условиях и генетике семей (раздел 3.6). С течением времени в семьях может увеличиваться количество паразитов и патогенов, что может нарушить предположение о независимости данных о выживаемости семей по годам, однако семьи не показали снижения вероятности выживания с увеличением возраста.
2.4 Оценка средней продолжительности жизни диких семей
Я оценил среднюю продолжительность жизни семей, используя вероятности выживания семей. Для этого я просуммировал по всем возрастам произведение каждого возраста на вероятность смерти в этом возрасте и добавил 1 год, чтобы перевести возраст смерти в продолжительность жизни. (Вероятность гибели в каждом возрасте рассчитывалась путем умножения вероятности выживания семьи до этого возраста и вероятности гибели семьи в этом возрасте (см. Seeley 1978). Для оценки средней продолжительности жизни семей, переживших рискованный первый год (= сформировавшихся семей), были проведены расчеты, описанные выше, но с вероятностью выживания 1,00 для первого года.
2.5 Создание имитационных семей в дикой природе
В течение трех летних месяцев (2011 – 2013 гг.) было создано 35 имитаций диких семей с использованием естественных роев: 19 собраны на ветках, 15 пойманы в ульях-приманках в лесу и 1 найден в пустом улье. Каждый улей состоял из сплошной нижней доски, «деревянного экрана Varroa» (Dadant and Sons, Hamilton, IL), корпуса улья Лангстрота с 10 рамками, деревянной внутренней крышки и телескопической внешней крышки (рис. 1). Сетка Varroa использовалась для подсчета осыпи клещей. Семьи имели соты, как в естественном гнезде (Seeley and Morse 1976). В каждом улье было восемь рамок, заполненных рабочим расплодом, и две рамки без расплода (чтобы у семей было место для строительства трутневого расплода). Пустые рамки находились в позициях 2 и 9. Вход в улей представлял собой отверстие площадью 22 см2 — средний размер для естественных гнезд (Seeley and Morse 1976). Каждый улей располагался на солнечном месте, входом на юг, на высоте около 0,3 м от земли, и был накрыт хозяйственной сеткой с ячейками 1/2″ (13 мм) для защиты от мышей. Через неделю после помещения роя в улей я помечал его маток краской (маркер POSCA на водной основе, Mitsubishi Pencil Co.), чтобы отслеживать смену маток.

2.6 Осмотр смоделированных диких семей и статистический анализ
Для определения продолжительности жизни и состояния семьи я осматривал каждую имитированную дикую семью три раза в год: в начале мая, в конце июля и в конце сентября. При каждом осмотре я осматривал соты семьи и отмечал: (1) жива или мертва семья, (2) цвет окраски матки, (3) наличие или отсутствие восковой моли (Galleria mellonella), (4) наличие или отсутствие болезней расплода: известкового (возбудитель — Ascosphaera apis), американского (Paenabacillus larvae) или европейского (Streptococcus pluton) гнильца. Заболевший расплод определяли визуально по Хансену (1987). Матки, не имеющие меток, получали метки цветом, соответствующим году. После каждого осмотра под сеткой семьи устанавливали липкую доску с решеткой (Dadant and Sons, Hamilton, IL, USA) для определения количества клещей V. destructor, упавших на нее за 48 ч.
Для проверки различий между летом и зимой в вероятности выживания семей я объединил количество семей, выживших или погибших в каждый сезон (лето или зима) за все 6 лет, в таблицу с осями лето/зима, основатель/учрежденный, выжил/погиб (см. Приложение 2, табл. V). Эти данные анализировали отдельно для летних и зимних семей, проверяя однородность доли выживших семей между основателями и укоренившимися семьями с помощью точного теста Фишера. По указанным выше причинам я предполагал независимость выживаемости семей по годам.
3 Результаты
3.1 Места гнездования диких семей и выживаемость
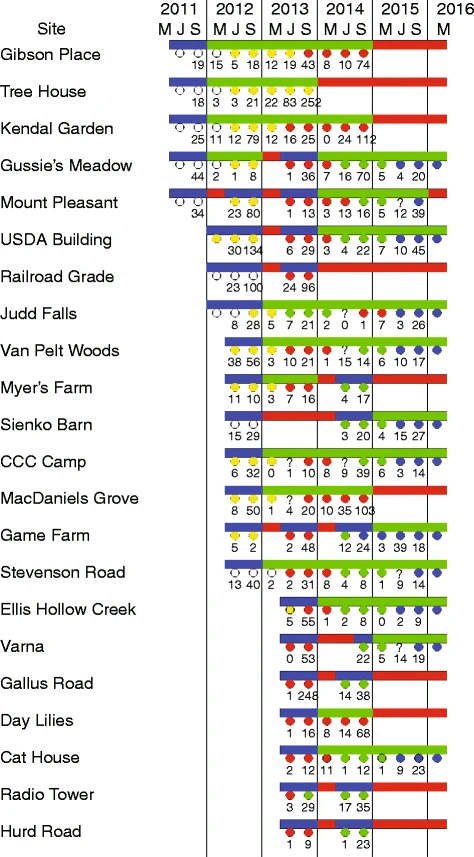
На рис. 2 для каждого из 33 гнездовых участков показано: (1) когда участок был добавлен в исследование, (2) какой тип участка, (3) сколько времени жила каждая семья и (4) когда каждый участок, освободившийся в результате гибели семьи, был вновь заселен. Большинство семей (61%) находились на деревьях, остальные (39%) — в зданиях. Пятьдесят три семьи занимали 33 участка.

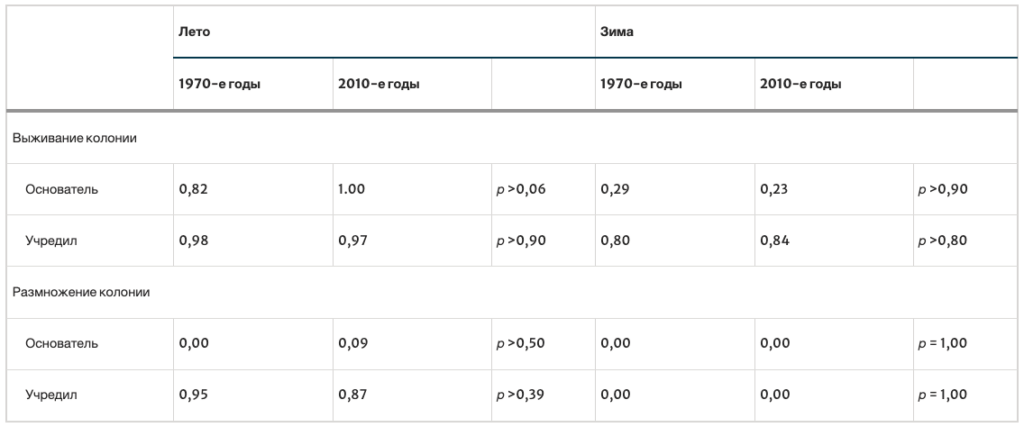
В таблице I приведены вероятности выживания семей для каждого года. За 7 лет было проведено 105 проверок выживаемости семей в летний период (26 семей-основателей и 79 укоренившихся семей) и 85 проверок выживаемости семей в зимний период (20 семей-основателей и 65 сформировавшихся семей). Как для семей-основателей, так и для сформировавшихся семей вероятность выживания летом была выше, чем зимой (для семей-основателей: p = 1,00 против 0,23, p < 0,0001; для сформировавшихся семей: p = 0,97 против 0,84, p < 0,0105). Из табл. I также видно, что семьи-основатели и сформировавшиеся семьи не различаются по вероятности выживания в летний период (p = 1,00 против 0,97, p < 0,58), но семьи-основатели по сравнению с сформировавшимися семьями имеют значительно меньшую вероятность выживания зимой (p = 0,23 против 0,84, P < 0,0001).
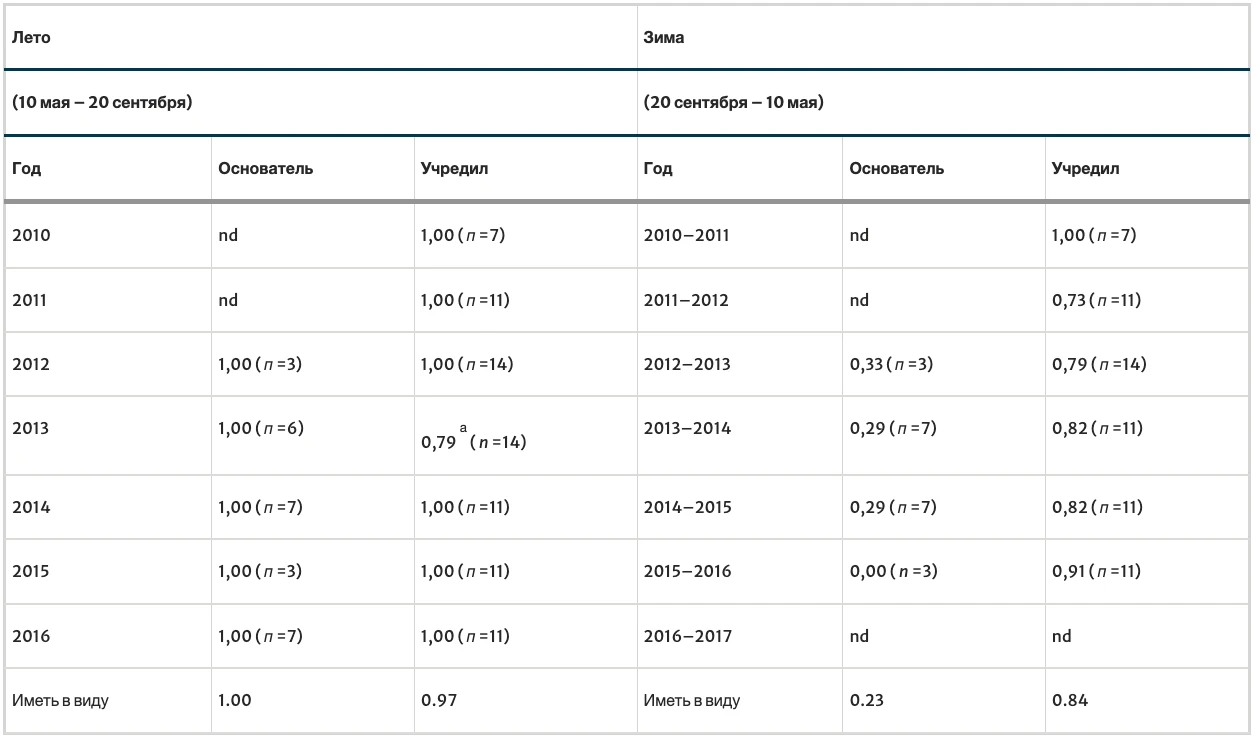
a Три семьи погибли: две — от медведя, разорвавшего гнезда в стенах хижины, и одна — от урагана, повалившего дерево.
3.2 Продолжительность жизни диких семей
Для оценки средней продолжительности жизни семей использовались средние вероятности выживания семей в летний и зимний периоды. Используя p = (1,00 × 0,23) = 0,23 для вероятности выживания в первый год (как семьи-основательницы) и p = (0,97 × 0,84) = 0,81 для вероятности выживания в последующие годы (как сформировавшейся семьи), я оцениваю среднюю продолжительность жизни семей в целом (семей-основательниц и сформировавшихся семей) в 2,2 года. Аналогичный расчет, но только для сформировавшихся семей, дает оценку средней продолжительности жизни семьи в 6,2 года.
3.3 Повторное занятие гнездовых участков дикими семьями
На рис. 2 показано, что в результате майских проверок был выявлен 31 случай гибели семьи за зиму. В результате гибели семьи образовалось свободное гнездовое место, оснащенное сотами и, возможно, мёдом и пыльцой. Многие из них были вновь заняты (21 из 31, 67%), причем нередко быстро (16 повторных заселений произошло в первое лето, когда участок был свободен, 3 — во второе лето и 2 — в третье).
3.4 Моделирование выживаемости диких семей
На рис. 3 для каждого из 22 ульев показано, когда он был добавлен в исследование, как долго каждая семья была жива в своем улье и как быстро каждый улей был вновь заселен после гибели семьи. В общей сложности за пять лет в этих ульях побывало 35 роев.
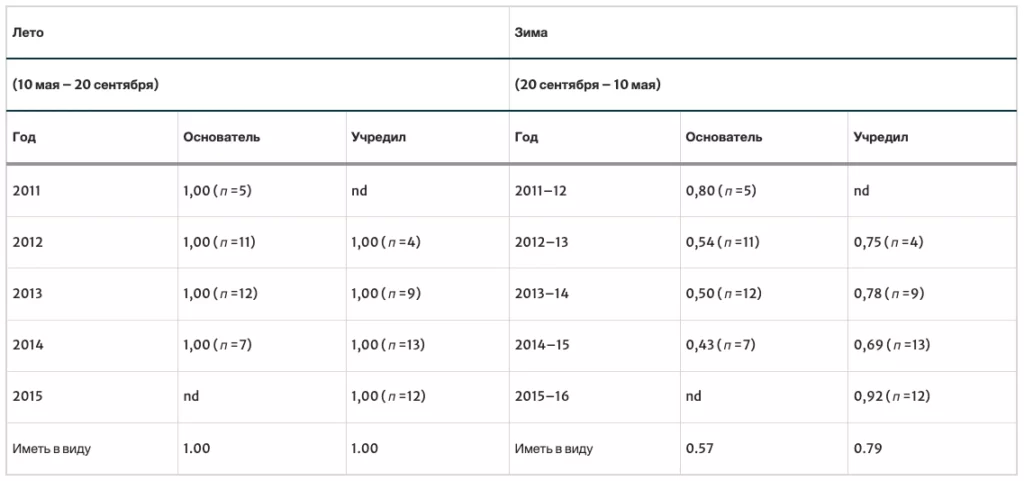
В табл. II приведены вероятности выживания семей для каждого года исследования. За 6 лет было проведено 73 проверки выживаемости семей в летний период (35 семей-основателей и 38 сформировавшихся семей) и 73 проверки выживаемости семей в зимний период (35 семей-основателей и 38 сформировавшихся семей). Вероятность выживания семей-основателей и сформировавшихсясемей летом была выше, чем зимой (семьи-основатели: p = 1,00 против 0,57, p < 0,0001; сформировавшихся семьи: p = 1,00 против 0,79, p < 0,006). Семьи двух типов не различались по вероятности выживания в летний период (p = 1,00 vs. 1,00, p = 1,00), но семьи-основатели по сравнению с сформировавшимися семьями имели более низкую вероятность выживания в зимний период (p = 0,57 vs. 0,79, p < 0,0764).
nd нет данных
3.5 Моделирование продолжительности жизни диких семей
Средние вероятности выживания семей в летний и зимний периоды использовались для оценки средней продолжительности жизни моделируемых диких семей. Используя p = (1,00 × 0,57) = 0,57 для вероятности выживания в первый год (как семья-основатель) и p = (1,00 × 0,79) = 0,79 для вероятности выживания в последующие годы (как сформировавшаяся семья), я оцениваю среднюю продолжительность жизни семьи в целом (основателя и сформировавшейся семьи) в 3,2 года. Проведя аналогичные расчеты, но только для сформировавшихся семей, я оцениваю среднюю продолжительность жизни семьи в 5,3 года.
3.6 Оборот маток в смоделированных колониях диких животных
На рис. 3 показано, что в течение 38 летних колоний проводилось наблюдение за укоренившимися семьями, и в 33 случаях была найдена немаркированная царица, которая затем была помечена новым цветом. Годовая вероятность смены маток в этих семьях составила 33/38 = 0,87.
3.7 Учет клещей в смоделированных семьях дикой природы
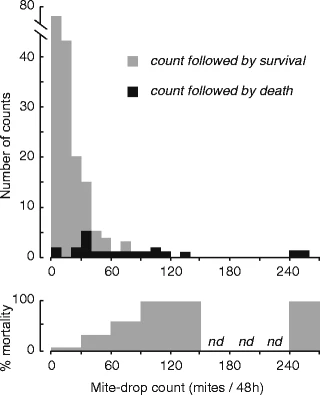
На рис. 3 показано 177 случаев падения клещей в 35 смоделированных семьях дикой природы. На рис. 4 показано распределение этих подсчетов. Большинство семей (141 из 177 = 80%) были низкими (0-29 клещей/48 ч), но некоторые были высокими. Колонии с числом клещей менее 30 клещей/48 ч имели практически нулевую смертность, но колонии с числом клещей более 30 клещей/48 ч имели более высокую смертность, достигавшую 100% при числе клещей 90+ клещей/48 ч.
На рис. 3 также показана зависимость между сменой цариц (роением) и количеством клещей в конце лета. В сентябре количество клещей в семьях с роением (23 ± 17 клещей/48 ч, n=33) значительно ниже, чем в семьях без роения (122 ± 75 клещей/48 ч, n=5) (t-тест; t(36) = 6,92, p < 0,0001).
3.8 Болезни и восковая моль в смоделированных семьях дикой природы
Частота встречаемости заболеваний, за исключением варроа и связанных с ним вирусов, была низкой. Был обнаружен только мелкий бругиоз, причем только в 4 из 35 семей и только в 6 из 197 проверок. Восковая моль не была обнаружена даже в ульях, простоявших без работы одно (n=9) или два (n=2) лета.
4 Обсуждение
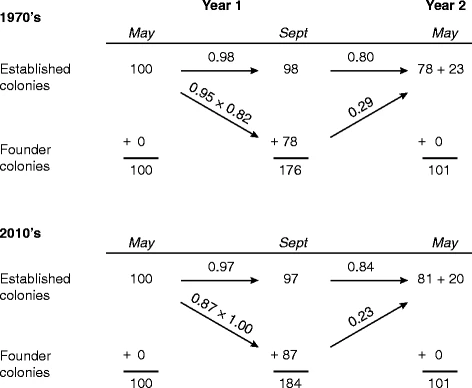
Основной целью данного исследования было выяснить, изменились ли особенности жизненного цикла семей диких пчел, обитающих в окрестностях Итаки (штат Нью-Йорк), после того как им пришлось вкладывать средства в иммунологическую, поведенческую и другие виды защиты от V. destructor и связанных с ним вирусов. В табл. III обобщены результаты, представленные здесь за 2010-2016 годы, и результаты, представленные ранее (Seeley 1978) за 1974-1977 годы. Эти два набора результатов поразительно схожи, несмотря на то, что они были получены примерно за 20 лет до этого и примерно за 20 лет после этого. 20 лет до и 20 лет после появления V. destructor. Оба набора результатов показывают, что большинство вновь основанных семей и укоренившихся семей выживают летом, но только около 26% семей-основателей и около 82% укоренившихся семей выживают зимой. Оба набора результатов также показывают, что семьи-основатели редко меняют маток/производят рои, но что устоявшиеся семьи, скорее всего, будут это делать (p = 0,96 для 1970-х гг. и p = 0,87 для 2010-х гг.). Кроме того, результаты 1970-х и 2010-х годов дают практически одинаковые оценки средней продолжительности жизни семей: 2,1 и 2,2 года для всех колоний и 5,6 и 6,2 года для укоренившихся колоний. Очевидно, что колонии, обитающие в окрестностях Итаки, в 2010-х годах имеют практически тот же набор характеристик жизненного цикла, что и в 1970-х: медленное развитие, большая продолжительность жизни после образования колонии и малое, но дорогостоящее потомство.
Модель динамики популяций 1970-х и 2010-х годов (рис. 5) предсказывает стабильную численность колоний для обоих периодов. Очевидно, что сохранение популяции диких колоний в прошлом и в настоящее время отражает активное размножение укоренившихся колоний, что приводит к почти двукратному увеличению популяции в летний период, а затем высокую смертность вновь образовавшихся колоний и низкую смертность укоренившихся колоний, что приводит к сокращению популяции в зимний период примерно вдвое.
Как объяснить тот факт, что дикие колонии 1970-х и 2010-х годов имеют практически одинаковый набор признаков жизненного цикла, несмотря на то, что дикие колонии 2010-х годов вынуждены вкладывать средства в защиту от интродуцированного паразита V. destructor и связанных с ним вирусов? Теория жизненной истории предсказывает, что затраты на защиту колонии компенсируются другими компонентами ее жизнеспособности, такими как скорость роста, выживаемость и размножение (Minchella 1985; Sheldon and Verhulst 1996; Schmid-Hempel 2003). Поэтому более значительные инвестиции в защиту колонии в 2010-х годах по сравнению с 1970-ми должны приводить к замедлению роста и ослаблению размножения, а возможно, и к снижению выживаемости.
Одно из возможных объяснений отсутствия изменений в признаках жизненного цикла заключается в том, что штаммы V. destructor (и/или переносимые ими вирусы) в диких колониях не были вирулентными, поэтому колониям не требовалась дорогостоящая защита от клещей и вирусов. Однако очевидно, что многие из имитированных диких колоний имели вирулентных клещей. Все колонии, в которых за лето не произошла смена королевы (роение?), к сентябрю имели смертельно высокую численность клещей (см. данные по колониям Gibson Place, Tree House, Kendal Garden, MacDaniels Grove и Day Lilies, рис. 3). Более того, все колонии с числом клещей более 100 (рис. 4) погибли, что позволяет предположить, что штаммы вируса деформации крыльев (DWV) в колониях были высоковирулентными, поскольку именно высокие титры вируса приводят к гибели колоний (Martin et al. 2012). Однако в недавней работе (Mordecai et al. 2015) сообщается, что авирулентный вариант DWV существует в некоторых местах в Великобритании, поэтому, возможно, авирулентный вариант DWV также существует в некоторых диких колониях около Итаки, штат Нью-Йорк. Эти колонии находятся на большом расстоянии друг от друга (Seeley et al. 2015), поэтому распространение V. destructor и DWV между колониями может происходить скорее путем вертикальной передачи (от родительской колонии к потомству, путем роения), чем путем горизонтальной передачи (между неродственными колониями; путем дрейфа, грабежа и фуражировки; см. Peck et al. 2016). Если это так, то, возможно, клещи и вирусы в этих диких колониях эволюционируют в сторону авирулентности (Ewald 1994; Fries and Camazine 2001), но представляется очевидным, что они еще не достигли этого уровня.
Наиболее вероятное объяснение отсутствия изменений в признаках жизненного цикла между 1970 и 2010 годами заключается в том, что дикие колонии обладают эффективными, но недорогими средствами защиты от V. destructor и связанных с ним вирусов. Мы знаем, что колонии, живущие в дикой природе, занимают небольшие гнездовые полости и часто роятся, и что эти черты диких колоний обеспечивают им устойчивость к Varroa (Loftus et al. 2016). Мы также знаем, что в результате эксперимента по оставлению колоний на Готланде в Швеции (Fries et al. 2003) выжившие колонии были гораздо меньше (и более склонны к роению?), чем исходные колонии (Locke and Fries 2011; Locke 2016). Кроме того, в настоящем исследовании было обнаружено, что колонии, в которых произошла смена царицы (вероятно, в результате роения), закончили лето с гораздо меньшим числом клещей, чем колонии, в которых смена царицы не произошла (в среднем 23 против 122 клещей/48 ч). Механизмы, благодаря которым маленькие и более роевые колонии обладают большей устойчивостью к V. destructor, до конца не изучены, но, вероятно, наличие относительно небольшого количества расплода, особенно трутней, помогает контролировать клещей. Также вероятно, что частое роение помогает бороться с клещами, поскольку во время роения вывозится около 35% клещей колонии; рабочие переносят около 50% взрослых клещей (Fuchs 1985), и около 70% рабочих покидают колонию во время первого роения (Rangel and Seeley 2012). Кроме того, роение временно лишает клещей кукольного расплода, а его отсутствие нарушает размножение клещей и повышает их подверженность воздействию клещегрызущих пчел (Hunt et al. 2016).
Поэтому, возможно, некоторые особенности биологии колоний медоносных пчел, живущих в дикой природе, такие как малый размер колонии и частое роение, обеспечивают им хорошую защиту от V. destructor, поэтому им не нужно эволюционировать дорогостоящие новые средства защиты от клещей и связанных с ними вирусов. Однако может быть и так, что диким колониям пришлось выработать новые средства защиты от V. destructor, включая гигиеническое поведение и поведение груминга, но эти новые средства защиты не требуют больших затрат. Наша следующая цель — определить набор защитных механизмов колонии, которые позволяют этой популяции диких колоний сохраняться, поскольку это позволит найти естественное, не требующее лечения решение проблемы V. destructor.
Ссылки
- Эвальд, П.В. (1994) Эволюция инфекционных заболеваний. Oxford University Press, Oxford.
- Fell, R.D., Ambrose, J.T., Burgett, D.M., DeJong, D., Morse, R.A., Seeley, T.D. (1977) Seasonal cycle of swarming in honey bees (Apis mellifera L.). J. Apic. Res. 16, 170-173
Fries, I., Camazine, S. (2001) Последствия горизонтальной и вертикальной передачи патогенов для эпидемиологии медоносных пчел. Apidologie 32, 199-214- Fries, I., Hansen, H., Imdorf, A., Rosenkranz, P. (2003) Роение у медоносных пчел (Apis mellifera) и развитие популяции Varroa destructor в Швеции. Apidologie 34, 389-397
- Fries, I., Imdorf, A., Rosenkranz, P. (2006) Выживание зараженных клещом (Varroa destructor) колоний медоносных пчел (Apis mellifera) в условиях скандинавского климата. Apidologie 37, 564-570
- Fuchs, S. (1985) Untersuchungen zur quantitativen Abschätzung des Befalls von Bienenvölkern mit Varroa jacobsoni OUD. und zur Verteilung des Parasitenim Bienenvolk. Apidologie 16, 343-368
- Hansen, H. (1987) Honey bee brood diseases. Wicwas Press, Ithaca, New York
- Hunt, G., Given, J.K., Tsuruda, J.M., Andino, G.K. (2016) Breeding mite-biting bees to control Varroa. Культура пчел 8, 41-47
- Korpela, S., Aarhus, A., Fries, I., Hansen, H. (1992) Varroa jacobsoni Oud. в холодном климате: рост популяции, зимняя смертность и влияние на выживание колоний медоносных пчел. J. Apic. Res. 31, 157-164
- Le Conte, Y., de Vaublanc, G., Crauser, D., Jeanne, F., Rousselle, J.-C., Bécard, J.-M. (2007) Колонии медоносных пчел, пережившие Varroa destructor. Apidologie 38, 566-572
- Locke, B. (2016) Естественные популяции медоносных пчел Apis mellifera, выжившие после нашествия клещей Varroa. Apidologie 47, 467-482
- Locke, B., Fries, I. (2011) Характеристика колоний медоносных пчел (Apis mellifera) в Швеции, переживших заражение Varroa destructor. Apidologie 42, 533-542
- Loftus, J.C., Smith, M.L., Seeley, T.D. (2016) How honey bee colonies survive in the wild: testing the importance of small nests and frequent swarming. PLoS ONE 11, e0150362. doi:10.1371/journal.pone.0150362
- Martin, S.J., Highfield, A.C., Brettell, L., Villalobo, E.M., Budge, G.E., et al. (2012) Global honey bee viral landscape altered by a parasitic mite. Science 336, 1304-1306
- Михеев, А.С., Тин, М.М.Я., Арора, Дж., Сили, Т.Д. (2015) Музейные образцы свидетельствуют о быстрой эволюции диких медоносных пчел, подвергшихся воздействию нового паразита. Nature Comm. 6, 7991. doi 10.1038/ncomms8991
- Minchella, D.J. (1985) Вариации жизненного цикла хозяина в ответ на паразитирование. Паразитология 90, 205-216.
- Mordecai, G.J., Brettell, L.E., Martin, S.J., Dixon, D., Jones, I.M., Schroeder, D.C. (2015) Superinfection exclusion and the long-term survival of honey bees in Varroa-infested colonies.
- Moritz, R.F.A., Kraus, B., Kryger, B., Crewe, R.M. (2007) Размер популяций диких медоносных пчел (Apis mellifera) и его значение для сохранения медоносных пчел. J. Insect Conserv. 11, 391-39
- Peck, D.M., Smith, D.L., Seeley, T.D. (2016) Клещи Varroa destructor могут ловко перебираться с цветов на кормящихся медоносных пчел. PLoS ONE 11(12): e0167798
- Potts, S.G., Biesmeijer, J.C., Kremen, C., Neumann, P., Schweiger, O., Kuning, W.E. (2010) Global pollinator declines: trends, impacts, and drivers. Trends Ecol. Evol. 25, 345-353
- Rangel, J., Seeley, T.D. (2012) Деление колонии у медоносных пчел: размер и значение фракции роя. Insectes Soc. 59, 453-462
- Schmid-Hempel, P. (2003) Вариации в иммунной защите как вопрос эволюционной экологии. Proc. R. Soc. Lond. B 270, 357-366
- Seeley, T.D. (1978) Life history strategy of the honey bee, Apis mellifera. Oecologia 32, 109-118
- Сили, Т.Д. (2007) Медоносные пчелы леса Арнот: популяция одичавших колоний, сохраняющихся с Varroa destructor на северо-востоке США. Apidologie 38, 19-29
- Сили, Т.Д. (2016) Вслед за дикими пчелами. Ремесло и наука охоты на пчел. Princeton University Press, Princeton
- Сили, Т.Д. Морс, Р.А. (1976) Естественное гнездо медоносной пчелы (Apis mellifera). Insectes Soc. 23, 495-512
- Seeley, T.D., Tarpy, D.R., Griffin, S.R., Carcione, A., Delaney, D.A. (2015) A survivor population of wild colonies of European honeybees in the northeastern United States: investigating its genetic structure. Apidologie 46, 654-666
- Sheldon, B.C., Verhulst, S. (1996) Экологическая иммунология: дорогостоящая защита от паразитов